THE THEORETICAL PREMISES OF THE ICDR METHOD
The Role of Carbon Dioxide in the Vital Processes of the organism (Chemical and Biochemical Aspects).
1. Forms of Carbon Dioxide
It is well known from the results of many investigations that carbon dioxide in the organism of Man and animals is not only a component of buffer systems and a regulator of respiration but is also a factor in the control of cardiac activity and of the vascular, hormonal, nervous and digestive systems.
At present, it can be said that carbon dioxide takes part in the control of many of the most important physical functions in the organisms of humans and animals.
When carbon dioxide is mentioned, often only one of its forms is implied, that is, the gaseous CO2 or carbon dioxide gas. When that is the case, what is being overlooked is that five different forms of carbon dioxide can be found in an organism:
- dissolved carbon dioxide gas (dCO2);
- carbonic acid (H2CO3);
- bicarbonates (HCO3– );
- carbonates (CO3)2–; {adapted}
- carbamates (R – NH COOH); {adapted}
{The following equilibrium equations adapted from an equilibrium diagram}
CO2 <=> pCO2
pCO2 + H2O <=> H2CO3
pCO2 + H2O <=> HCO3– + H+
pCO2 + R_NH2 <=> R_N_(H2+)_COO–
pCO2 + OH– <=> HCO3–
H2CO3 <=> HCO3– + H+
HCO3– <=> (CO3)2– + H+
2. The Role of Carbon Dioxide in the Respiratory Process.
The main source of carbon dioxide in an organism is indigenous carbon dioxide. The intensity of CO2 formation in tissues in the processes of metabolism can vary in a considerable range depending on various factors, but the normal CO2 concentration in blood and other biological fluids in a organism is not subject to any major fluctuations. As a rule, it is maintained at a strictly, defined level and any excess is immediately removed. This removal is ensured by the function of a complex system of mechanisms controlling the concentration. Of these the most important is the processes of neutralisation and rapid removal of excess CO2 from the tissues into the environment. This mechanism can be understood as follows.
The CO2 which forms inside cells penetrates into the extracellular fluid and then into the bloodstream. The mechanism of CO2 transfer through cellular membranes has not yet been fully established. All that is known is that membranes are more permeable to CO2 and H2CO3 than to HCO3–. As the CO2 permeates the interstitial fluid and then enters into the bloodstream most of it is transformed into HCO3–. This process is catalysed by the enzyme, carboanhydrase, (CA) of erythrocytes:
CO2 + H2O <=(KA)=> H2CO3 {+} H+ + HCO3
As a consequence of this process the pH inside erythrocytes begins to fall. At certain pH values the glyoxaline imidazole group of histidine and the amino group of valine, both situated on the chains of hemoglobin, begin to incorporate H+ ions which leads to the acquisition of a positive (+) charge. This charge stabilises the salt bridges which have the function of preserving the desoxystructure of hemoglobin. Apart from this process, another process of replacement of Cl ions by carbon dioxide takes place in the net of electrobathic interactions, thus leading to an increase in the stability of the quaternary structure of hemoglobin. Further, the carbon dioxide influences the binding of DPH and binds to -amino groups, which also stabilises the desoxystructure of hemoglobin (the Verigo-Bohr effect).
On account of the difference in concentrations of oxygen in erythrocytes and tissues, the oxygen diffuses into the tissues. As a consequence of the cooperative O2-binding effect, the separation of succeeding oxygen molecules is made less difficult.
The summary results of the two processes are the maintenance of a practically constant pH, which aids the diffusion of oxygen and the retention within the erythrocytes of K+ ions, which were neutralised by HbO2 earlier and are subsequently neutralised by the forming HCO3– ions.
As a result, the major part of CO2 which diffused out of the tissues into the erythrocytes transfers from capillaries into veins in the form of HCO3– ions contained in erythrocytes.
Due to this isoacidic shift, the ratio of HCO3– in erythrocytes (e) to HCO3– in plasma (p) is altered as a result of the HCO3– increase in erythrocytes. No longer is the ratio of HCO3–e/CL–e maintained at a level equal to the ratio of HCO3–p/CL–p which had been established through the manifestation of the Gibbs-Donnan effect.
The tendency of HCO3– to leave the cells rises and the resulting loss of HCO3– ions is compensated by Cl– ions entering from the plasma until a new equilibrium is achieved. The final result of this process is the situation where a major portion of the total amount of CO2 which had entered into the erythrocytes in the capillaries, is then transferred in the form of HCO3– into venous plasma.
It should be noted that the transformation of HbO2 into HbH+ and the resulting buffer action of this transformation does not influence the osmotic pressure due to the hemoglobin of cells because the amount of hemoglobin does not change, but the total effect of the isoacidic and chloride shifts does result in an increase in the total quantity of anions and this does raise the effective osmotic pressure in cells. As a consequence, water is redistributed between the cells and the plasma.
This process takes a reverse direction in the lungs. Because of the difference in pressures oxygen penetrates into erythrocytes and forms acid HHbO2. As a result of its dissociation H+ ions are formed which react with HCO3– forming H2CO3. H2CO3 splits into CO2 and H2O under the influence of carboanhydrase:
H2CO3 => H2O + CO2
CO2 and H2O are released into alveolar spaces.
The release of CO2 leads to a shift in Bohr’s reaction to the left side which results in a normalisation of pH and facilitates the binding of oxygen to hemoglobin.
While this process takes place in the lungs, the processes of carbonate splitting and of the transfer of proteins into a free state proceed at the same time.
Bicarbonate, which has entered the blood reacts with carbonic acid and forms the pair HCO3–/H2CO3 which is a component of the main buffer system in extracellular fluid. The need for the existence of precisely such a buffer is determined by many reasons:
- There is considerably more HCO3– than other buffer compound in the extracellular fluid:
- The intake of CO2 is not limited:
- The physiological mechanisms which maintain the normal pH value of extracellular fluid do so by regulating the concentrations either of HCO3– or CO2. Van Slyke and then Warburg found that there is a linear relationship between pH,, dCO2 and HCO3– in blood described by the Henderson-Hasselbach equation of:
pH = 6.1 + lg([HCO3–]/[pCO2]) - the HCO3/H2CO3 buffer system functions in concert with the Hb system, as described above.
As in all buffer systems, the pH does not depend on the absolute concentrations of the buffer components but on their ratios (Tables 1, 2, 3 from the book: Robinson J.R. Grounds for the Regulation of an Acid-Alkaline Equilibrium. Moscow, Meditsina, 1969).
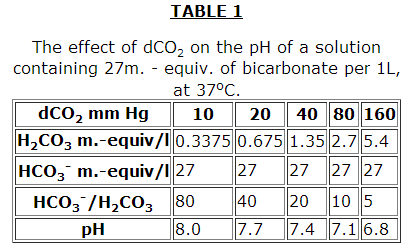
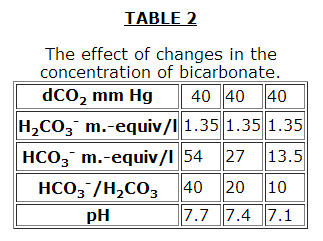
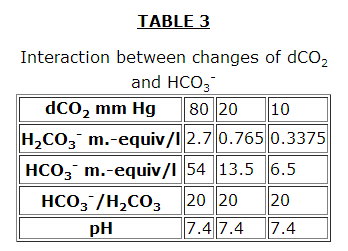
However, it should be noted in Table 3 that although the pH is kept constant, the buffer capacity of the plasma is decreased. The effectiveness of the HCO3– buffer system increase considerably in the presence of erythrocytes.
The described combination of buffer properties, where one of the components is a gas and where automatic self regulation is achieved by means of intracellular Hb, lead to a remarkably stable pH being maintained in blood plasma.
In addition, the organism uses two other stabilising systems, the respiratory system and the kidneys which correspondingly control (H2CO3) and (HCO3–), and thereby produce additional possibilities for the maintenance of a constant pH in extracellular fluid.
3. Control of CO2, HCO3– and H+ Levels in the Tissues of Animals.
As distinct from HCO3– (fixed anion concentration), (H2CO3) is determined solely by one parameter: the partial pressure of CO2 in the gaseous mixture in equilibrium with extracellular fluid as present in alveolar air. This pressure in turn depends on the rate at which the CO2 passing out of the blood in the lungs is diluted by atmospheric air and so it depends on the depth and the frequency of breathing. The character of respiration is regulated by a respiration centre in the nervous system; apparently this centre is sensitive to pH and dCO2 in the extracellular fluid. When the pH of the extracellular fluid falls below a normal value because of a decrease in HCO3–, respiration is stimulated which leads to a lowering of alveolar dCO2 and consequently of extracellular (H2CO3).
This restores the ratio of (HCO3–)/(H2CO3) to the normal value of 20: 1 and leads to the return of pH to the normal value. The resultant fall in CO2 pressure in plasma acts in an opposite direction upon the regulating nervous centre, so the compensation would never be completed if only this regulatory mechanism was functioning.
At high levels of plasma pH the frequency of respiration falls and alveolar dCO2 and therefore (H2CO3) in plasma rise which then shifts the pH down to a normal value. In this case full compensation is not achieved because the elevated plasma (H2CO3) acting on the respiratory centre produces an effect opposite to that of increased pH. if the frequency of respiration is considerably lowered, a decrease in dO2 will stimulate an increase in respiratory activity.
At the time when the respiratory mechanism compensates for deteriorations of acid-alkaline equilibrium by means of H2CO3 regulation in extracellular fluid, kidneys participate in pH control by regulating HCO3–. Kidneys counteract the decrease in extracellular pH, caused by the increase of alveolar dCO2 or by the decrease in HCO3–, by the two ways available to them, namely, by liberation of H+ in the form either of undissociated acid or NH4+.
Renal compensation, under the conditions which could bring about an increase in extracellular pHi is accomplished by means of a decrease in HCO3– of extracellular fluid.
There is evidence that simultaneously with these mechanisms which compensate for extracellular pH fluctuations the “potassium-sodium pump” also comes into action. Muscular tissues, epithelial cells of renal ducts, and, possibly, all other cells have an ion-exchange mechanism which carries out exchange of either K+ or H+ or both of these ions for Na+through the cell membrane. Due to this exchange, the cell content can participate in maintaining extracellular pH.
With increase in extracellular HCO3–, Na+ enters the cells in exchange for H+ and K+. Protons react with extracellular HCO3– and the CO2 formed is removed with exhaled air, K+ is removed with urine together with the equivalent amount of HCO3–. With the decrease in pH, Na+ leaves the cell and H+, K+ ions enter it.
By direction of shift of active reaction of blood all changes of acid-alkaline balance are divided into acidosis and alkalosis, and due to the reason causing changes in pH of blood they are divided into metabolic and respiratory.
4. Shifts of Acid-Alkaline Equilibrium Respiratory Acidosis: This is characterised by the following factors: dCO2> normal rangel; pH< 7.36; (HCO3–)..normal range or above it. These states are observed, with the inhibition effect on the respiratory centre by morphine, during inhalation of gaseous mixtures containing a high percentage of carbon dioxide, in severe cases of pneumonia, oedema and emphysema, during spontaneous arrest of respiration, with asthma and suffocation.
Respiratory Alkalosis: Factors:dCO27.44; (HCO3–) … normal range or slightly lower than the normal range. This state is developed during hyperventilation of lungs, with severe inflammation of internal sex organs, hypoxia and impaired blood circulation. With prolonged respiratory alkalosis an increase in lactic acid, which forms due to the incompletely oxidised pyruvate, that is, in the absence of sufficient oxygen concentrations in tissues, is constantly observed.
Metabolic Acidosis: Factors: PH [ normal range; (HCO3–)[ normal range; dCO2 [normal range. Metabolic acidosis develops in cases of origination of excessive amounts of non-volatile acids in extracellular fluid which shows itself:
- with the intake of food containing a large amount of non-volatile acids;
- with the formation of large amounts of organic acids (lactic, acetoacetic, pyruvic, etc.) during oxygen insufficiency in tissues;
- due to deterioration of the renal function in elimination of hydrogen ions;
- with a relative excess of non-volatile acids, due to the loss of bases during vomiting, diarrhoea, fistula, etc.
Metabolic Alkalosis: Factors: increase in PH; (HCO3–)> normal range; dCO2 >normal range. Metabolic alkalosis develops in the case of loss of non-volatile acids or in the case of loss of K+ (this results in excessive elimination of H+ by the kidneys).
Extensive distribution of the above mentioned states of the organism of animals and man became a main reason for numerous investigations carried out since the end of the 19th century up to the present time. These have been devoted to the study of characteristic properties of metabolism in tissues during various conditions of alkalosis and acidosis. However, approaches to the study of this problem and explanation of obtained results, for a long time, in the majority of cases, were based solely on the role of change in concentration of hydrogen ions and hardly any attention has been paid to the important metabolic meaning of change of various forms of carbon dioxide in tissues under these conditions.
In recent years interest in clarification of the metabolic meaning of carbon dioxide has increased. It has been established that CO2 takes part in regulation of numerous key points of metabolism and respective physiological functions of an organism. In particular, it has been shown that the intensity of biosynthetic processes of basic organic components of animal tissue is directly proportional to the intensity of carbon dioxide fixation processes (reactions of carboxylation).
5. Reactions of Carboxylation
Only recently has it become known that processes of carbon dioxide fixation lie at the basis of all important metaboIic and primarily biosynthetic processes in tissues. Up to thepresent a number of reports has been published on these topics. Summing up accumulated data, it is possible to conclude that a great number of reactions exists in tissues of an organism, which depending on the source of used energy can be divided into three groups:
a) reactions taking place at the expense of energy of ATP(these are reactions of carboxylation of propionyl – CoA, pyruvate, acetyl CoA, -methyl-crotonyl-CoA, geranyl-CoA, urea and formation of carbomylphosphate). Here fixation of carbon dioxide is preceded by its activation, whic is accomplished by its relationship with ATP. An important role in this case is assigned to biotin a prosthetic group of almost all carboxylases catalysing ATP – dependent reactions of carboxylation.
These process of carboxylation using biotin can be represented by the scheme: {Modified after scanning}
b) reactions proceeding at the expense of the energy of the reduced forms of pyridinenucleotides (this is a reaction of carboxylation of alpha-ketoglutarate, pyruvate, ribulose-5-phosphate, serine, glutamate, acetate);
c) reactions not requiring energy from outside (5-aminoimidazole-ribotidecarboxylase, phosphoenylpyruvate-carboxylase, and phosphoenolpyruvate carboxy – transphosphorylase reactions belong to this group of reactions.)
Analysing the data on CO2 fixation in all three types of reactions, a number of factors influencing this process can be indicated. Of these, the main factors are the following:
- variations of concentrations and magnitudes of the relationship of basic forms of carbon dioxide in tissues (HCO3– and dCO2);
- change in concentrations of certain ions of metals which are activators and inhibitors of carboxylases;
- change in concentrations of substrates subjected to carboxylation;
- change in the degree of provision of cells by energy sources as well as by a number of other metabolit-allosteric effectors of carboxylases;
- change in concentrations of carboxylasaes coenzymes in tissue.
The first is the most universal from all indicated ways of the effect on the carbon dioxide fixation because in all reactions of carboxylation dCO2 and HCO3– are substrates. Concentration of the substrate in enzymatic reactions can even play a certain role in intensity of the process of its transformation. It was shown in many experiments under the conditions of normal pH of incubative medium the intensity of carbon dioxide fixation in tissues IN VITRO increased as its concentration increased. Thus, it can be concluded that the intensity of carboxylation reactions in many instances is determined by concentration and by magnitude of dCO2 and HCO3– ratios in tissues. Under the conditions of constant pH the possibility exists for considerable increase in intensity of the carboxylation processes by increasing carbon dioxide concentration up to a certain level in tissues.
It has been established in many investigations that metabolic importance of carbon dioxide is not limited by the above mentioned suggestions. It was found that when changing carbon dioxide level in tissues, the activity of a wide variety of enzymes of glycolysis, tricarboxylic cycle and nitrogenous metabolism changed and the formation of some hormones, etc.increased.
6. Role of Carbon Dioxide and Regulation of Reactions of Tricarboxylic Acids Cycle (TAC).
{Diagram of the Tricarboxylic Acids Cycle omitted}
Considering the role of carbon dioxide in the regulation of reactions of tricarboxylic cycle the following should be noted. Many biochemists elaborated the erroneous idea that the cycle of tricarboxylic acids was only a mechanism of oxidation of acetyl-coenzyme A. For this reason, such names as “energy-mill”, “kettle”, etc. were secured for TAC, and almost all biochemistry textbooks and even in many monographs, TAC was presented as a sequence of reactions with the process occurring only in a clockwise direction.
In consequence often the question is raised: What is the role of carbon dioxide in providing normal functioning of the cycle, in the sense that in one cycle in the reactions of carboxylation two molecules of CO2 are formed, but only one CO2 is used in the formation of oxaloacetate (carboxylation of pyruvate). Further, because during a cycle the oxaloacetate is again regenerated, then it is sufficient that once a molecule of oxaloacetate has been formed it will constantly maintain the cycle in a functioning state. An impression can be had that the carbon dioxide is not practically required for the functioning of the cycle. Such suggestions are fundamentally erroneous. The error of these notions is as follows.
1) Almost all reactions of TAC are reversible. They proceed in both directions in a cell. At reverse direction (in an anti-clockwise direction) the carbon of carbon dioxide will not be liberated but used in the reactions of carboxylation for build-up of a carbon frame of a number of organic compounds, this means then that carbon dioxide should come from outside. In this case, the concentration of bicarbonates and CO2 in a cell will greatly influence the rate of these reactions and as a consequence the state and function of TAC.
2) TAC plays an important role by providing the basis for oxidising and biosynthetic processes where it serves as a mechanism for generating numerous substrates used in the synthesis of many amino acids and of the nitrogenous bases of purine and pyrimidine nucleotides for carbohydrates and lipids.
3) A considerable number of TAC metabolites are used in the process of transmembrane transport of various anions and other substances. There is a view that about 40% of the TAC metabolic pool is consistently removed and used in various biosynthetic processes. If this view is right, then normal functioning of TAC is possible only when the means for continuous formation of its metabolites are not disrupted.
Because 4 of the 9 metabolites (oxaloacetate, malate, succinate, isocitrate) are formed as a result of the carboxylation of pyruvate and, phosphoenolpyruvate, propionyl-coenzyme A, alpha-ketoglutarate, functioning of the cycle is impossible without constant inflow of bicarbonate and carbon dioxide.
Summarising the results of various experiments in this respect (Adler et al., Ostberg et al.), it can be said that the activation of TAC is determined largely by the magnitude of pH of biological fluids and by the level of bicarbonates and CO2 contained. At the same time, under the conditions of constant pH of biological fluids, the activity of TAC also increases if their concentration of carbon dioxide increases. The experimental results of Berri, Kun and Werner are among convincing confirmations of the importance of carbon dioxide and the processes of its fixation in the regulation of oxidation processes of organic substances in the cell. These experiments showed that during incubation for 40 minutes of the isolated liver cells of rats in a medium containing bicarbonate, the concentration of malate increased approximately 7 times in comparison with the concentration in a medium without bicarbonate (phosphate buffer), although pH of the media were the same.
Thus it can be concluded that carbon dioxide, being one of the basic final products of metabolic reactions of TAC, appears as one of its powerful regulators. The results of the latest investigations confirm that the mechanism of regulatory effect of carbon dioxide on the functioning of TAC is not only in the fact that it is one of the components of carboxylation or decarboxylation reactions. The fact is that the activity of certain enzymes, which are not related to the processes of carboxylation and decarboxylation, can be substantially changed at-constant pH of the medium depending on the changes in HCO3– and CO2 concentrations in the medium (even in the ranges of physiological fluctuations).
Apart from this the obtained data show that the level of carbon dioxide determines not only the intensity of functioning of the indicated reactions, but also influences the level of biosynthetic processes in an organism, which also are in continuous dependence on the functioning of tricarbonic cycle processes of metabolism of lipids, carbohydrates, proteins, nucleotides and nucleic acids belong to such processes.
7. The Role of Carbon Dioxide in Processes of Biosynthesis
Biosynthesis of Amino Acids: Synthesis of such amino acids as glycine, serine, citrulline, glutamic and aspartic acids and respectively of their amides, glutamine and asparagine, almost fully depends on the intensity of carboxylation of the corresponding substrates:

Biosynthesis of Nitrogenous Bases of Nucleotides
Biosynthesis of purine nucleotides: The first important data from biosynthesis of purine bases were obtained by Beaukenen who fed animals with various labelled precursors and determined spots of inclusion of the labelled atoms in the purine ring. These experiments were carried out on birds whose nitrogen is liberated mainly in the form of uric acid, which represents a derivative of purine. It was found that two nitrogen atoms N (3,9) of the purine ring were derived from the amide group of glutamine, the third nitrogen atom (1) was derived from aspartic acid and finally, the last (7) had its origin in glycine.

The fourth and fifth carbon atoms also had their origin from glycine. Thus, glycine molecule gave three atoms enclosed in a frame. The second and eighth carbon atoms originated from formate, and the sixth carbon atom came from CO2.

The second carbon atom of pyrimidine is formed only at the expense of carbon of the CO2 molecule. Thus, this suggests that without carbon dioxide, biosynthesis of such compounds is not possible. Considering that in biosynthesis of nitrogenous bases amino acids such as aspartate, glycine and glutamine participate, whose formation depends on the corresponding reactions of carboxylation, it can be said that biosynthesis of these bases, and also of nucleic acids is directly dependant upon the carboxylation reactions.
Biosynthesis of Lipids: When studying the destiny of the labelled carbon atoms of the CO2 molecule in liver extracts which synthesised fatty acids with a long acetate chain, Wakephil and other authors observed that carbon atoms were found in one of the carboxyl groups of malonyl-CoA.
Malonyl-CoA is formed during carboxylation of acetyl-CoA
COOH | CH3CO.CoA + CO2 + ATP <== Mg2+ ==> CH2 + ADP + P | CO.CoA
and is an immediate precursor of two carbon units in synthesis of fatty acids.
Biosynthesis of Carbohydrates:
The majority of stages of well studied biosynthetic course of the glucose formation from pyruvate is catalysed by enzymes of glycolytic cycle, and, thus, they represent reversal of the reactions forming in the process of glycolysis. However, three irreversible stages exist in the normal glycolytic course which cannot be used during conversion of pyruvate into glucose; the biosynthesis proceeds by by-passing these stages using alternative reactions which favour the synthesis thermodynamically.
One of them is the change of pyruvate into phosphoenolpyruvate. Usually this reaction does not proceed by means of direct application of pyruvatekinase reaction. Phosphorylation of pyruvate is achieved using an indirect succession of reactions:
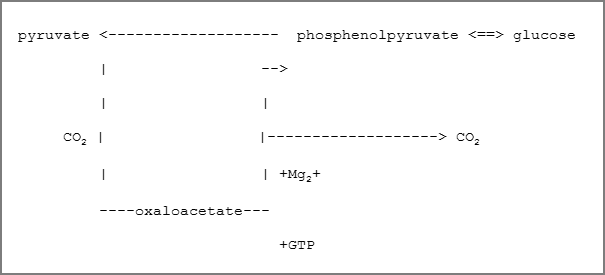
It is very important to emphasise that carbon dioxide fulfils the catalytic function both in lipogenesis and gluconeogenesis because its carbon in these reactions is not directly incorporated into either fatty acids or phosphoenolpyruvate. However, the importance of these reactions is high because they are limiting (reactions) in the indicated processes.
Taking into account the participation of carbon dioxide in all basic biosynthetic and oxymethylic processes in a cell, it can be proved that without CO2 life is absolutely impossible.
THE ROLE OF CARBON DIOXIDE IN PROCESSES OF ACTIVITY OF AN ORGANISM AT THE LEVEL OF SYSTEMS
1. The Role of Carbon Dioxide in Regulation of Activity of the Nervous System
It has been known for a long time that an increase in the level of CO2 in inhaled air has a narcotic effect on the organism of humans and animals. An inhalation of gaseous mixture containing 10% CO2 produces a dazed state in people and at higher concentration they even lose consciousness. Under the conditions of increase in bicarbonate and CO2concentration the synthesis of acetycholine in the nervous tissue intensifies and transmembrane potential of nervous cells changes because of the transfer of negatively charged hydrocarbonate ions through conjugated membrane according to the concentration gradient: from cell to medium or from intermitochondrial space to cytoplasm, and also according to the electrical potential. In this case, a discharge of the transmembrane and of energy potential of the cells occurs. To maintain its stability, consumption of ATP is required. When CO2 is added to the medium (addition – of carbonate and hydrcarbonate) the gradient of HCO3– concentration will decrease and if pH of the medium is higher than pK of carbonic acid, the. transmembrane gradient HCO3– will decrease more considerably than the gradient CO2 and H2CO3. This means that the rate of transfer of HCO3– ions is reduced according to the concentration gradient, the discharge rate of the electric potential of the transmembrane and the use of ATP for supply of energy to the cell membranes.
Apart from participation in the function of the nervous system, carbon dioxide plays an important role in the activity of the regulation of hormonal, cardiovascular and digestive systems in humans and animals.
2. The Role of Carbon Dioxide in the Regulation of Hormonal System
At the present time data are available on the change in intensity of functioning of the individual endocrine glands in animals when the level of bicarbonates and CO2 in their tissues changes. A number of authors obtained interesting results on the effect of change in the hormonal status of an organism on the character of carbohydrate metabolism depending on the concentration of HCO3–, CO2 and H+. For example, Kaplan et al. observed the stimulation of parathormone formation in sheep after the intravenous injection of NaHCO3solution. The other investigators noticed the rise in epinephrine formation in dogs and rabbits during conditions of acidosis.
It has also been reported that carbon dioxide increases sensitivity to insulin, through the processes of galactose transfer through cell membranes of myocardium in rats and that extracellular bicarbonate and CO2 have a stimulating effect on the secretion of insulin by cells of pancreas in rats.
3. The Role of Carbon Dioxide in the Regulation of Cardiovascular System.
It has been found that under the conditions of hypercapnia increase in blood volume per minute, reduction in frequency of heart contractions, some increase in blood pressure and change in tone of cerebral, coronary and peripheral vessels take place.
It is of interest that the indicated effect of CO2 on the vascular tone can be twofold: the reflex when hypercapnia increases the excitability of the vasometer centre and brings about stenosis of peripheral vessels and the local effect when the increase in concentration of CO2 in lumen of vessels causes dilation of vessels. Thus, the final effect of CO2 will be determined by the reaction of these two processes. In particular, it has been shown that during hypercapnia the vessels of the brain, heart and intestines are dilated and the vessels of skeletal muscles are considerably constricted. In addition, in the experiments using volunteers, whose levels of bicarbonate and CO2 were increased in the blood of one of the humeral arteries (without pH change), a considerable vasolidating effect of the carbon dioxide was observed.
4. The Role of Carbon Dioxide in the regulation of the Digestive System
The data from the effect of the level of bicarbonate and CO2 in tissues on the activity of the system of digestive glands of animals and humans are very interesting. It was shown that the concentration of CO2 in a medium had an important significance in the regulation of hydrochloric acid formation (linear relationship). The linear relationship is also observed between the intensity of gastric secretion and the level of CO2 in contents of the duodenum in man.
5. The Role of Carbon Dioxide in the Regulation of the Immune System
The data which confirm the suggestion that the change in carbon dioxide content affects the activity of the immune system have been accumulated in the literature. Some of them are illustrated as examples.
In 1980, several authors in the article “Differential Reactions for Stimulation of Lymphocytes by Mitogens in Healthy Persons and in Patients with Lesch-Nyahan Syndrome: the Effect of Bicarbonate Buffer System” noted the rise in the proliferative reaction of lymphocytes which were deficient in hypoxanthine – guanine – phosphoribotransferase in response to the action of the polyclonal activators in the case where lymphocytes were cultivated in an atmosphere containing 5% CO2 in the bicarbonate buffer system, the proliferation of lymphocytes in patients with Lesch-Nyhan syndrome was damaged under the influence of some stimulators of the T-cells (PHA (phytohemagglutinin), ConA (concanavalin A), streptokinase-streptodornase). The authors of this paper assume that the obtained data indicate a decisive role of DNA denovo biosynthesis in the mitogenic stimulation of lymphocytes.
Additionally, the increase in total carbon dioxide (HCO3 and CO20 content in blood of chickens reportedly stimulated biosynthesis of antiplague by 12-21% in comparison with control, this is explained by the increase in biosynthesis of protein and biopolymers, also including antibodies (I. N. Zhuravski et al).
At the same time a great deal of information is available about the role of pH in the process of binding of the antibody with the antigen. A deviation of the pH to the side from a certain optimum results in a decrease in the affinity and therefore in the weakening of the immune reaction. Further, reports have been published on the change in specificity of antibodies as a result of the change in pH.
Thus, for example, Arepalli S. Rao with other authors during the study of precipitation reaction of human macroglobulin IgM-WEA using polysaccharides Klebsiella K30 and K21 showed that IgM-WEA, which precipitates at pH 7 with polysaccharide K30 and does not react with polysaccharide K21, was losing its specificity when pH changed down to 4-4.5. At pH 4-4.5, IgM-WEA reacted with the polysaccharide K21 in the same manner as with the polysaccharide K30 at pH 7, but at pH 4-4.5 the reaction of IgM-WEA with polysaccharide K30 was considerably weaker than with the polysaccharide K21.
Thus, it can be said that, at least, three factors which depend on the CO2 concentration produce a substantial effect on the activity of immune system.
THE FOUNDATIONS OF BUTEYKO’S THEORY
1. The Purpose of the Invention
The significance of K.P. Buteyko’s invention resides in the fact that at deep respiration, an excessive removal of CO2 takes place from an organism, resulting in the change of homeostasis which is partially neutralised by various compensating mechanisms. The developing disruption of homeostasis, accumulating for a long time (in fetus through mother’s respiration) brings about the disruption of the above-mentioned biosynthetic and other processes. The state of health of people who suffer from shortage of carbon dioxide is impaired by defensive reactions of the organism displayed in the form of disruption of the processes of respiration and hemodynamics. However, it should be noted that if these defensive reactions which aggravate the health state of sick persons had not existed, these people would perish much faster.
2. The First Statement of the Theory
The deep respiration during which the rate of CO2 exhalation from the organism somewhat exceeds the rate of its accumulation in the tissues, develops into a respiratory alkalosis characterised by the decrease in CO2 and the increase in pH.
The respiratory alkalosis is accompanied, as a rule, by the state of oxygen deficiency which develops as a result of the inhibition of Verigo-Bohr effect, this brings about an occurrence of surplus of incompletely oxidised products of metabolism in the blood and therefore a development of the metabolic acidosis, which somehow compensates the deviations of pH caused by the respiratory alkalosis. However, it should be noted that this compensation is not fully adequate. As a result of these processes in the organism the following changes develop:
1) a shift occurs in the electrolytic composition of the extracellular fluid;
2) a shift occurs in pH of the extracellular fluid;
3) a shift of pH occurs in the cells as a consequence of the participation of cells in the reaction of pH normalisation of the extracellular fluid;
4) as a result of the renal activity in the elimination of HCO3. which is connected with a tendency of the organism to normalise the ph, a depletion of the buffer capacity occurs in plasma;
5) owing to the chanqe in ratio HCO3– cell/HCO3– plasma the diffusion rate of HCO3– changes from cell to blood plasma;
6) efficiency of the process is disrupted
7) disruption of the processes of synthesis: amino acids, nitrogenous bases, fats and carbohydrates.
The effect of all these changes is as follows:
1) a disruption of the formation: of proteins and peptides, of nucleic acids, lipids and carbohydrates;
2) a disruption in the work of enzymes originating as a result of the change in pH, a change in the carboxylation intensity;
3) a disruption in the metabolizable energy of the organism (a decrease in the formation of ATP).
Because of the above-mentioned disruptions the changes originate in various systems of an organism. At the decrease in dCO2 and [H+] which develop as a consequence of deep respiration, the respiratory centre adapts to these concentrations with the result that an accommodation of deep respiration occurs. Later when CO2 concentration becomes sufficiently low, the organism will feel acute shortage of O2 and will start to react, not on the excess CO2, but on the shortage of O2 by depressing the respiration. As a result of this process, a positive reverse bond originates which will bring about an increase in the intensity of work of the system and in our case will even more depress the respiration up to the advance of an untimely catastrophe.
It is quite natural to assume the presence of certain defensive mechanisms which prevent excessive exhalation of CO2 from the organism. Such mechanisms can be : stenosis of blood vessels, change in heart activity and constriction of respiratory pathways.
It is assumed that a choice of the defence method depends on the individual characteristics of the organism as well as on the intensity and breadth of processes leading to the depression of respiration. Because of this in a number of cases the constriction of lumen of vessels in a certain organ preferably takes place against the background of less noticeable constrictions of lumen of the remaining vessels and respiratory pathways of the organism, and in a number of cases a radical change in the respiration and (or) circulation takes place.
3. Comparison of Theoretical Conclusions with the Experimental Material
Let us consider how these conclusions of Buteyko’s agree with the facts. In the book: ” Introduction to Clinical Biochemistry ” edited by Prof. I.I.Ivanov, Corresponding member of the- Academy of Medical Sci. of the U.S.S.R. (1969), it is quoted that patients having mitral defects of the heart show a clear relationship between the degree of development of the pathological process and the state of myocardial metabolism. During IV – V stages of the disease (according to the classification by Bakulaev) a decrease in the use of oxygen and succinatede-hydrogenase activity is observed in the muscle of auricula of the left atrium and the concentration of creatine phosphate decreases; additionally the intensity of glycolysis rises and the content of inorganic phosphate increases.
Also there is indicated that in 1963 Karsanov found a decrease in the total amount of actomyosin in the extracts from myocardium of persons who had died from chronically congested heart deficiency which originated as a result of defects, hypertony in systemic and pulmonary blood circulation and cardiosclerosis. The greater the decrease in the content of actomyosin in the extracts from from various heart sections, the higher the deficiency of the heart contracting function. In particular, in the composite mitral defect the actomyosin decreases preferably in the extracts from the atrium, right ventricle and right atrium. In the heart diseases resulting mainly in the deficiency of left ventricle, the decrease in content of actomyosin in the extracts from the left ventricle prevailed over its decreas in the extracts from the right ventricle.
F.Z.Meerson and T.Ya Zeyats (1960) assume that the disruption of constricting function of myocardium during chronic heart deficiency can be connected with the deterioration of protein synthesis in the myocardium and the exhaustion of plastic reserve of cells.
In rabbits with stenosis of the aorta, F.Z- Meerson and G.P. Ramenskaya (1960) observed a considerable decrease in DNA content in myocardium.
It is well known that one of the most characteristic pathological processes in the nervous tissue which develop as a primary damage or are accompanying other functional disorders of the nervous system, is the demyelinisation process. The nature of this process consists in the disintegration of structural, well-regulated on the molecular level, protein-lipid complex which forms myelin sheaths of grey matter nerves. It is thought that the demyelinisation process is connected with the change in the enzymic activity of lipid metabolism.
R.N.Phil’kevich in the article “Change of Protein Metabolism in Patients with Dermatosis” (Collection of Sci. Reports of the Central Committee Viral Institute, MZ U.S.S.R., 1978, No.22) indicates the presence of disproteinaceous shifts during the investigations of the protein fractions of blood serum using electrophoresis.
V.S.Shchelkunov et al. in the article “The Effect of Unloading Diet Therapy on the Protein Metabolism, Dynamics of the Exchange of Circulating Blood and Extracellular Fluid in Acute Pancreatitis” (Acute Pancreatitis. Biochemical Aspects. Leningrad, 1978) showed that considerable protein-volemic disruptions developed in patients with the edematose form of acute pancreatitis.
There is a great amount of data on the disruptions of protein metabolism in ulcer disease and atherosclerosis, etc.
There is little need to mention the enormous amount of literature published in connection with the metabolic change of fatty acids and the decrease in absorption of O2 in the heart during ischemia.
I think, it is worth noting the article by I.I.Zhuravasky et al. “The Effect of the Change in Carbon Dioxide (HCO3– and CO2) Level in the Blood on the Biosynthesis of Antibodies in Chickens” (Reports of the U.S.S.R- Academy of Sci., 1981, NoB), in which particularly it is confirmed that the origination of metabolic acidosis in chickens inhibits the formation of antibodies by 12.7 – 32.6% in comparison with the control.
In the article “Cromolin and Bronchial Spasm Induced by Deep Respiration”, Pneumonologie, 1975, 153 (1), Messerlich et al. write about the development of bronchial spasm caused by the deep respiration. Kilham H. et al. in the article “Running, Walking, Hyperventilation Causing Asthma in Children” draw a conclusion that the hyperventilation brings about asthma in children.
Finally, in order to have a full picture, I would like to mention the article of Chr. Marinov “The Change and Some Mechanisms of the Disruption in Acid-Alkaline Equilibrium in Acquired Heart Defects”, where it is, noted that in the study of 76 patients with the acquired heart defects and heart deficiency of various degree, the presence of respiratory alkalosis accompanied by metabolic acidosis was established.
It can be said that the obtained data are in full agreement with the conclusions made on the basis of Buteyko’s theory. Namely, in various diseases both the changes in pH just in the direction predicted by the theory, and the changes in biosynthesis of polymers as well as in the absorption of oxygen by tissues have been noted.
Thus, the theory being developed on the basis of the internal logic, results in a number of deductions which do not contradict the independently obtained experimental data. This means that the developed theory has the right to be considered suitable for the explanation and structuring of the existing experimental material in reference to the process of origin of the diseases.
4. The Second Statement of the Theory
According to the first statement of Buteyko’s theory two components can be indicated in the concept “disease”:
1) a disease as the change of functions in the organism owing to the deficiency in CO2;
2) a disease as the defensive reaction of the organism to the removal CO2.
Because the second component follows from the first component, according to Buteyko’s opinion, it is sufficient to eliminate the disease connected with the deficiency of CO2. The effect of this will be the elimination of the disease connected with the defensive reaction of the organism to the removal of CO2.
The reverse process of the reaction is possible because all the discussed biochemical ways are reversible, if not directly, then via the stages.
Thus, Buteyko’s discovery makes it possible at least to stop the course of the disease and improve a patient’s state of health by intensifying various metabolic reactions and other processes and thus to reverse the course of the disease up to its complete regression, restoring the initial status of the organism.
It should be noted that at the present time in a majority of cases the main efforts of doctors are directed to treat a disease connected with the defensive reaction of the organism to the deficiency of CO2 (this represents a search and use of all available spasmolytics, activators of heart activity,, etc.) . According to Buteyko’s idea this makes the basic disease connected with the deficiency of CO2 worse, and never leads to curing the second disease, since the symptoms of the basic disease being Just somehow suppressed in one place can start developing in another place.
5. The Method for Intentional Cessation of Deep Respiration
For elimination of the basic disease K.P Buteyko offered the method for intentional cessation of deep respiration (ICDR).
On decrease of deep respiration less oxygen enters the lungs, CO2 is accumulated and the respiratory centre becomes adapted to its high concentrations. As a consequence of intensified Verigo-Bohr effect the oxygen from the blood enters more actively into the tissues, this leads to better oxidation of the substrates and therefore to the decrease in the concentration of incompletely oxidised products. The increase, in [H+] caused by ICDR method is eliminated by the function of kidneys, which, on the one hand, intensify the removal of H+ ions and on the other hand, slow down the removal of HCO3–. As a result of this process, the concentration of non-volatile acids decreases and the buffer capacity of plasma increases. This together with the restoration of all above mentioned processes culminates in the destruction of the basic disease.
6. Conclusion
The main statements of Buteyko’s theory are based on the enormous amount of experimental material on the role of carbon dioxide in the processes of an organism activity and on the concepts of reversibility of biochemical processes, involving the regressive changes in the development of diseases, which were repeatedly verified during the approbations in Moscow and Leningrad.
BIBLIOGRAPHY
- Buteyko K.P. Oscillogtaphs and Hypertonia. Is “Enhanced” Breathing Beneficial? Izobretatell I Ratsionalizator. 1962. No.5.
- Buteyko K.P. Shurgaya Sh.I.Functional Diagnosis of Coronary Disease. Thesis from Symposium on Surgical Treatment of Coronary Disease, Moscow, 1962.
- Buteyko K.P., Demin D.V. Cross-correlational Analysis of Physiological Functions. Izv.Sib.Otd. AN SSSR. 1963, No.6. Ser.biol.med.nauk.Vypusk Issue) 2.
- Buteyko K.P., Odintsova M.P., Demin D.V. The Influence of Hyper and Hypoxaemia on the Tonus of Peripheral Vessels. Proceeding of the Second Siberia Scientific Conference of Therapists, Irkutsk, 1964.
- Buteyko K.P., Demin D.V., Odintsova M.P. The Application of Regressive Analysis for Differentiation of the Influence of Gaseous Components of the Arterial Blood on the Functional State of Fine Peripheral Arteries. Materialy Vtoroy sib. Nauchnoy Konf. Terapevtov. Irkutsk, 1964.
- Buteyko K.P., Demin D.V.,, Odintsova M.P. The Mathematical Analysis of Interreaction of Physiological tigation Factors. Physics-Mathematical Methods for Inves in Biology and Medicine. Materialy Pervoy Novosib- Conf., Novosibirsk/, 1965.
- Buteyko K.P., Demin Dove,, Odintsova M.P. The Relationship between the Ventilation of the Lungs and the Tone of Arterial Vessels in Patients with Hypertonic Disease and Stenocardia. Fiziologichni Zhurnal, 1965. Vol.II, No.5 (in Ukrrainian).
- Buteyko K.P., Demin D.V., Odintsova M.P. The Relationship between the Carbon Dioxide in Alveoles, Arterial Pressure and Cholesterol in Blood in Patients with Hypertonic Disease and Stenocardia. Materialy Tretley Sib. Nauchn. Conf. Terapevtov. irkutsk, 1965. (Proceedings of the Third Siberian Sci. Conference of Therapists).
- Buteyko K.P. The Cross-Correlational Technique of Analysis of Physiological Regulations. Materialy VII Vsesoyuzn. Conf. po Avtomat. Kontrolyu i Metodam Elektr. Izmereniy. Novosibirsk, 1965, (Proc. of the VII-Union Conference on Automatic Control and Methods for Electrical Measurements).
- Buteyko K.P. The Complex Investigations into Functional Systems in Biology and Medicine. (Reports of the Medical Electronics Section of the 9th Regional Sci.-Tech. Conference Dedicated to the Day of. Radio). Dokl. Sektsii Med. Elektroniki 9-oy Obl. Nauchn.-Tekhn. Conf. Posvyashchennoy Dnyu Radio, Novosibirsk, 1966.
- Buteyko K.P., Demin D.V., Odintsova M.P. The Relationship between the Partial Pressure in Alveolar Air and the Tone of Peripheral Arteries in Patients with Hypertonic Disease and Stenocardia. Zdravookhranenie Turkmenistana. 1966. No.2 (Public Health of Turkmenistan. 1966, No.2).
- Buteyko K.P., Demin D.V., Odintsova M.P. The Effect of Carbon Dioxide on the Coronary Efferent Arteries in Patients with Hypertonic Disease and Atherocardiosclerosis. Zdravookhranenie Kazakhranenie Kazakhstana. 1966. No. 6 (The Kazakhstan Public Health, 1966, No.6).
- Buteyko K.P., Demin D.V., Odintsova M.P. A Linear Model for Regulation of Vascular Tone by Gaseous Components of Arterial Blood. Dokl. 9-oy Obl. Nauchn.-Tekhn. Conf., Posvyashchennoy Dnyu Radio. Novosibirsk, 1966.
- Buteyko K.P. et al. The Influence of Intentional Regulation of Respiration on Some Physiological Functions at Altitude. Voprosy Aviatsionnoy Meditsiny (Problems of Aviation Medicine). Moscow, 1966.
- Buteyko K.P., Chasovskikh S.F.,, Demin D.V. An Indirect Determination of the Degree of Hypertension of Pulmonary Circulation According to the Data of Calibrated Phonocardiography. Materialy 4-oy Povolzhskoy Conf. Fiziologov, Biokhimikov i Farmakologov- Saratov, 1966. T.l. (Proceedings of the 4th Volga River Conference of Physiologists, Biochemists and Pharmacologists. Saratov, 1966, Vol.1).
- Buteyko K.P., Odintsova M.P., Demin D.V. The Influence of Hyper- and Hypoxaemia on the Tone of Arterial Vessels. Sovetskaya Meditsina. 1967. No.3 (Soviet Medicine, 1967. No3)-
- Murakhtanova Z.M., Buteyko K.P. et al. Concerning Characteristic of External Respiration in Patients with Scoliosis. Sbornik Rabot Novosib. Instituta Travmatologii i Ortopedii. Novosibirsk, 1967. (Collection of Reports of the Novosibirsk Institute of Traumatology and Orthopedics. Novosibirsk, 1967).
- Buteyko K.P. et al. The Use of Correlation Methods for Investigation of Cardiovascular System. Mat. Metody v Aviatsionnoy i Kosmicheskoy Meditsine. – Moscow, 1967 (Mathematical Methods in Aviation and Cosmic Medicine).
- Buteyko K.P., Demin D.V., Odintsova M.P. The Mutual Information of Heartrythm and Other Physiological Human Functions according to the Data of Cross-Correlative Analysis. Mat. Metody Analiza Sergechnogo Ritma. Moscow, 1968 (Mathematical Methods for Analysis of Heart Rythm. M., 1968).
- Buteyko K.P., Odintsova M.P., Nasonkina P.S. The Ventilation Test for Patients -with Bronchial Asthma. Vrachebnoe Delo, 1968, No.4 (Medical Treatment. 1968, No.4).
- Buteyko K.P., Odintsova M.P. Hyperventilation as One of the Reasons for the Spasm of Nonstriated Musculature of the Bronchi and Arterial Vessels. Materialy 4-oy Nauchn.-Praktich. Conf. po Vrachebn- Kontrolyu i Lechebnoy Fizkulture. Sverdlovsk, 1968 (Proceedings of the 4th Scientific-Practical Conference on Medical Control and Medical Physical Culture. Sverdlovsk, 1968).
- Buteyko K.P. Complex Methods for Investigation of Cardiovascular System and Respiration. Voprosy Funktsionallnoy Diagnostiki. Novosibirsk,, 1969 (Problems of Functional Diagnostics. Novosibirsk, 1969).
- Buteyko K.P. The Theory of CO2-Deficient Diseases of Civilisation as an Adaptation to the Evolution of Atmosphere. Kiberneticheskie Aspekty Adaptatsii Sistemy “Chelovek-Sredall. Tez. Seminars. Moscow, 1975 (Cybernetic Aspects of Adaptation of the System “Man-Environment”. Seminar Thesis, M., 1975).
- Buteyko K.P. Therapy of Hemohypocarbia. Patent No.1067640. Registered in the State Record of Inventions of the U.S.S.R., 15.9.1983.
- Buteyko K.P., Genina V.A. Concerning Pathogenesis of Asthma Attack during Physical Exercise. Non-medical Methods for Treatment of Patients with Bronchial Asthma. Tez. Dokl. Vsesoyuzn. Conf. Moscow, 1986 (Report Thesis from the All-Union Conference. M., 1986).
- Buteyko K.P., Genina V.A. The Comparison of Justification and Effectiveness of the Complex, Symptomatic and Monoetiological Principles of Treatment of Bronchial Asthma and Other Allergies. Ibid.
- Buteyko K.P., Genina V.A. The Theory of Discovery of Deep Respiration (Hypervention) as a Main Cause of Allergic,, Bronchial-vascular-spasmodic and Other Diseases of Civilisation. Ibid.
- Buteyko K.P., Genina V.A., Nasonkina N.S The Reactions of Sanogenesis in Medical Treatment Using the ISDR Method. Ibid.
- Putintsev E.V., Varlamova Z.A., Meshcheryakova L.R. Concerning Medical Physical Culture for-Bronchial Asthma. Sb.Materialov k Godichnoy Nauchnoy Sessii Instituta. Novokuznetsk, 1967 (Collection of Reports for Annual Sci. Session of the Institute. Novokuznetsk, 1967).
- Shelomova K.V. Intentional Normalisation of Respiration in the Complex of Medical Physical Culture for Bronchial Asthma. Proceedings of the 4th Sci. -Practical Conf. on Medical Control and Medical Physical Culture. Sverdlovsk, 1968 (as in 22).
- Panova L.A. Bronchial Asthma – a Model of Adaptation to the Changed Environmental Conditions. (As in 24.).
- Gavalov S.M., Genina V.A., Gavalova R.F. Intentional Regulation of Respiration in Complex Medical Treatment of Bronchial Asthma in Children. Saratov, 1976.
- Genina V.A., Glur-,henko I.R. The Construction of a Generalised Indicator of the Bronchial Asthma Condition. Methods for Physical Measurements of Parameters of the Ecological Systems and the Mathematical Processing of the Obtained Data. Novosibirsk, 1982.
- Genina V.A. The Role of Hyperventilation in the Pathogenesis of Bronchial Asthma and Its Medical Treatment by means of Reduction in Ventilation of the Lungs. Epidemiological Characteristics of Non-r.,pecific Diseases of the Lungs in Various Professional Groups. Novosibirsk, 1982.
- Genina V.A. et al. The Medical Treatment of Bronchial Asthma in Children Using the Method of Intentional Normalisation of Respiration (INR) according to the Data of the 1st MMI Children’s Clinic. Pediatriya, 1982, No.2 (Pediatrics, 1982, No.2).
- Genina V.A. The Technique for Elimination of Chronic Hyperventilation in the Treatment of Patients with Obstructive Syndrome. Medical Treatment of Non-specific Diseases of the Lungs.
